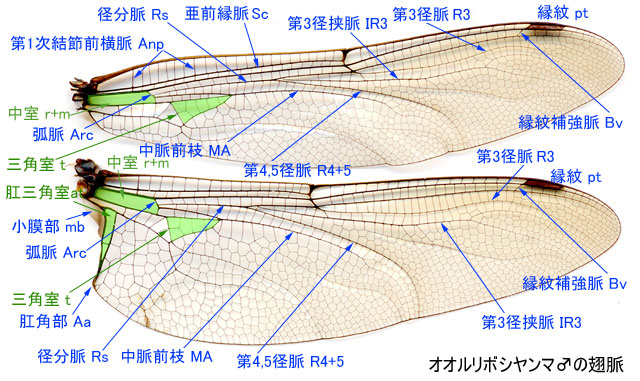
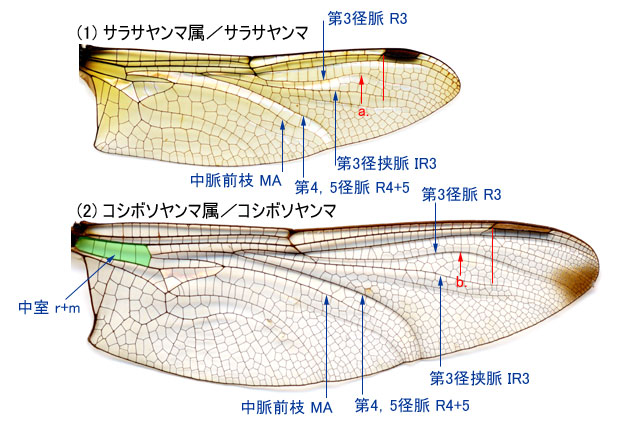
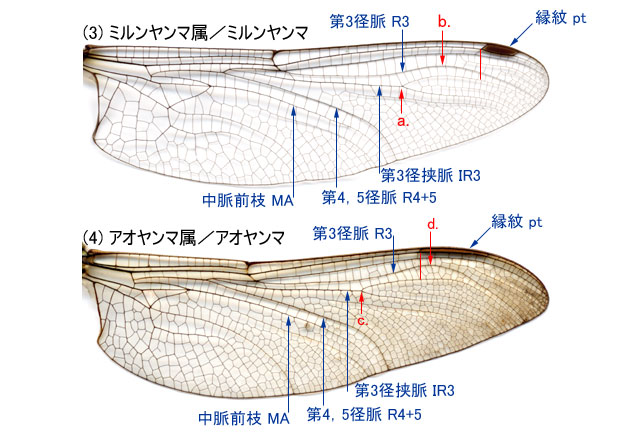
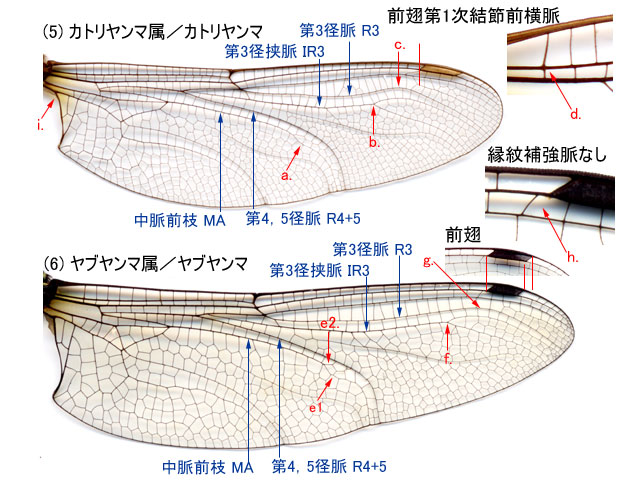

-
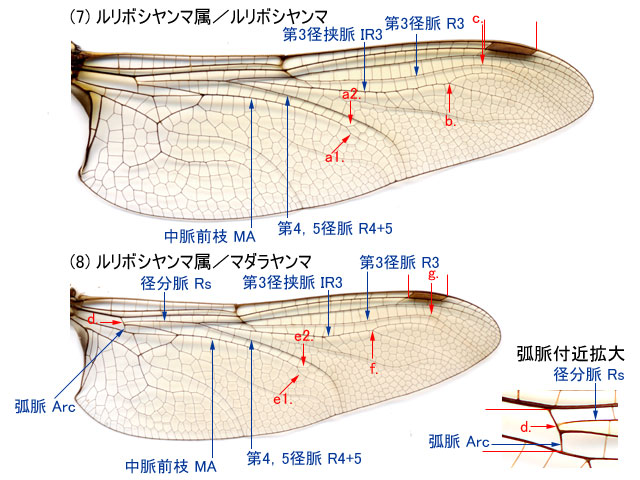
a.第3径脈 R3 は,縁紋内縁付近またはやや内側の位置で後方に曲がる(5c).前翅の第1結節前横脈 Anp は亜前縁脈 Sc とほぼ直角に交わる(5d).小膜部 mb がない(5i)・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・カトリヤンマ属
b.第3径脈 R3 は,縁紋内縁から外縁付近の位置で後方に曲がる(6g)(7c)(8g)(9e)(10i).前翅の第1結節前横脈 Anp は亜前縁脈 Sc と斜めに交わる.・・・・・・・・・・・・・2. -
a.縁紋補強脈 Bv がないかまたは不明瞭(6h).前翅の縁紋は後翅の縁紋より長い(6).・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ヤブヤンマ属・ヤブヤンマ
b.縁紋補強脈 Bv は明瞭.前翅の縁紋は後翅の縁紋とほぼ同じ長さ.・・・・・・・・・・・3. -
a.径分脈 Rs は弧脈 Arc の中点付近または少しだけ前方から分枝していて(8d),第3径脈 R3は,種によって縁紋付近の様々の位置で後方に曲がる.♂の肛角部は先端が鋭く尖る.・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ルリボシヤンマ属
b.径分脈 Rs は弧脈 Arc の中点よりかなり前方から分枝していて(9a),かつ,第3径脈 R3は,縁紋の外側端くらいの位置で急角度で後方に曲がる(9e)(10i).♂の肛角部は存在しない場合もある(10f).・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・4. -
a.第3径脈 R3 は,縁紋付近の位置で,特に縁紋に近づくように膨らむことがなく急角度で後方に曲がる(9e).♂の後翅には肛角部があり(9b)肛三角室がある.・・・・
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・トビイロヤンマ属
b.第3径脈 R3 は,縁紋の外側端付近の位置で,縁紋に近づくように膨らんで曲がる(10i).♂の後翅に肛角部がなく(10f)肛三角室もない.・・・・・・・・・・・・・ギンヤンマ属
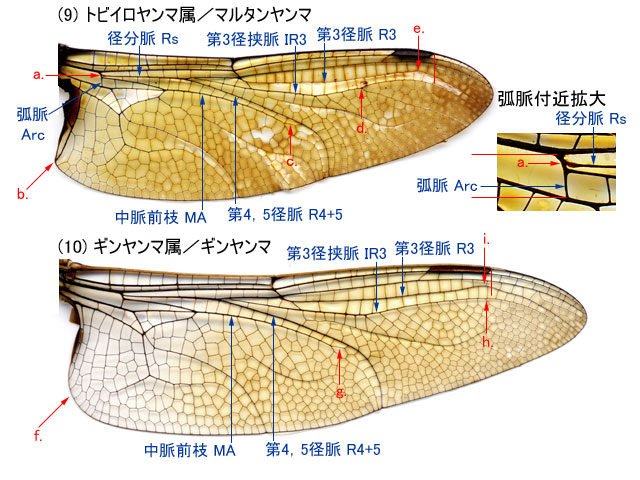
図7.マルタンヤンマ♂とギンヤンマ♂の後翅翅脈 (Aeshnini [3]).
中脈前枝 MA は,後方に曲がるあたりで-
第4,5径脈 R4+5 に合流するような横脈が顕著に表れる(c,g).
第3径挟脈 IR3 は先の方で分枝する.(d,h).
マルタンヤンマでは第3径脈 R3 は,縁紋外縁付近で後方に曲がる(e.).
ギンヤンマでは第3径脈 R3 は,縁紋外縁近くで縁紋の方に膨らんでから曲がる(i).
径分脈 Rs は弧脈 Arc のかなり前方で分枝する.
中脈前枝 MA は,後方に曲がるあたりで-
第4,5径脈 R4+5 に合流するような横脈が顕著に表れる(c,g).
第3径挟脈 IR3 は先の方で分枝する.(d,h).
マルタンヤンマでは第3径脈 R3 は,縁紋外縁付近で後方に曲がる(e.).
ギンヤンマでは第3径脈 R3 は,縁紋外縁近くで縁紋の方に膨らんでから曲がる(i).
径分脈 Rs は弧脈 Arc のかなり前方で分枝する.