トンボの生態学
6.成虫の飛行移動 目次
トンボの生態学 6.成虫の飛行移動
トンボの翅と飛行
トンボは,誰が言ったのかは分かりませんが,その名称が「飛ぶ棒」から来ているという説があるくらい,飛ぶことに特化した昆虫です.その翅の構造や巧みな飛行術は,航空力学の研究対象として,専門的な研究がいろいろとなされています.ここではそういったことには深入りせず,その生態的学的な意味について考えることを中心にしたいと考えています.
トンボには4枚の翅がありますが,はばたいて飛ぶときには,前翅と後翅の動き(位相)を少しずらせています(写真2左).特に空中の一点に止まって行う停止飛翔(ホバリング)を行う際には,ほぼ交互になるようにはばたきます(写真1左).つまり前翅を打ち下ろしたときには後翅が上がり,後翅を打ち下ろしたときには前翅が上がるという具合です.ただし翅の上下動は,体軸に直角ではなく,やや前方に打ち下ろしそのままやや後方に上がるような動きをしており,同時に翅を少しねじらせるような動きをしています(写真1左;写真2右).一方全くはばたかずに,風に乗るように滑空飛行(滑翔)をする場合もあります(写真1右).
写真1.停止飛翔するオナガサナエの翅のはばたきの動き(左)と,はばたかずに滑空飛翔するウスバキトンボ(右).
写真の上にマウスカーソルを置くとアニメーションが始まります.
写真2.均翅亜目のはばたき.ハグロトンボの直線飛行で後翅が少し下の位置にありずれている(左).ホソミイトトンボの直線飛行時のはばたき(右).
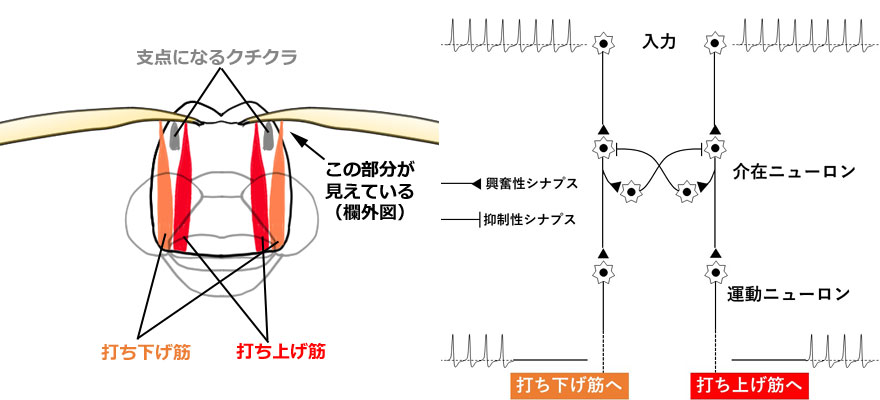
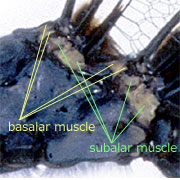
トンボの翅は横紋筋でできた筋肉によって動きます(Tillyard, 1917).トンボの翅を動かす筋肉は,神経のパルスと同期しており,1つのパルスで1回の収縮が起きます.このような筋肉を同期筋と呼びます.筋肉の一端は,支点となるクチクラの内側にある翅の基部,および外側の翅に直接結合していて(直接飛翔筋と呼ばれる),交互に収縮することで翅を動かします(図1左).トンボを捕まえて翅を背中側でたたむように持つと,そのつけ根の部分に,翅基部の外側に付いている,翅を打ち下ろすための筋肉(腱)を観察できます(右欄外図).Snodgrass (1935) によると,それらには basalar muscle, subalar muscle という名称がつけられており,前後翅にそれぞれ2本ずつあります.翅の筋肉付着部は,それぞれ肩板 humeral plate,腋板 axillary plate と呼ばれる堅いクチクラででできています.そしてこれら4本の筋肉の個別の動きによって翅の上下動やねじれの動き(内転・外転運動)など,複雑な動きを実現しています.
このような規則正しい連続的なはばたき運動を実現する中枢神経回路はバッタや脊椎動物でよく研究されており,中枢パターン発生器 (Central Pattern Generator;CPG) と呼ばれています.最近の高校生物の教科書にも載るぐらい一般的です.特に意識しないでも持続するリズミカルな運動をつかさどる神経回路には,この CPG が存在すると考えられています.CPG によって生み出される運動は,視覚・触覚などの感覚情報によって動きを調整できるようになっており,とっさの場合にその強さ,向きなどを変更できるようになっています.例えば歩行運動を考えてみると,あまりよい例ではありませんが,スマホを操作しながらでも歩けるのは,CPG によって自動的に歩行運動が生成されているからです.そして人影が視界に入ったとき,とっさによけることも可能です.
非常に大きな筋肉であり,力強く羽ばたくことができる.
図1.トンボの直接飛翔筋の模式図と,はばたきをつくりだす CPG の簡略化モデル(実際は感覚情報の入力などがありもっと複雑).
図の上にマウスカーソルを置くとアニメーションが始まります.
トンボのはばたきについてはまだ CPG の存在は確かめられていないようですが,おそらくよく似たしくみがあると思われます.そして風の強さ,向き,気温,視覚情報などの感覚情報を使って,微妙に飛び方を調整しているのでしょう.なお直接飛翔筋では,はばたきの回数はそれほど多くなく,例えば野外での計測で,Epitheca cynosura で43Hz や Anax junius で29Hz (May, 1991),実験条件下のギンヤンマで 26~29Hz (Azuma & Watanabe, 1988)など,せいぜい1秒間に数十回というところです*1.
飛行速度については,Tillyard (1917) が偶然の機会に計測した非常に極端な例があって,「Austrophlebia 属*2のトンボ(ヤンマ科)が,80~90ヤードの直線部分を3秒で飛んだ」と記しています.1ヤードは0.9144メートルですから,時速に直すと,なんと約 88-98km/h になります.彼は,これ以上のスピードを出せるトンボがいるとは思えないと述べています.計画的に記録された別の速い例では,Aeshna cyanea で 10m/s (時速 36km),A. junius で 7.5m/s (時速 27km) という計測値がありますが,これは最大速度であり普段はもっとゆっくりと飛んでいるということです(Rüppell, 1989).
はばたき飛行のエネルギー源はグルコースです.筋肉にグリコーゲンとして蓄えられており,これを分解してグルコースにして利用しています.筋繊維(筋細胞)内にはたくさんのミトコンドリアがあって,短時間に大量のエネルギーを生み出します.ただ長時間飛行の際にはグルコースではなく脂質を代謝するような変換が起き(Kallapur & George, 1973),より長時間エネルギーを生み出すことができるようになっています.Kallapur (1985) は,ウスバキトンボについて,糖類と脂肪の蓄積量から推定して,最大8時間ぐらい「はばたき飛行」ができると述べています.
沖ノ鳥島はフィリピン海に浮かぶ岩礁ですが,ここでウスバキトンボが採集され,私のもとへ送られてきたことがあります(欄外写真).ここから最寄りの繁殖可能な陸地,例えばグアム島までは最短距離でも600kmはあります.ウスバキトンボは,多分時速88-98kmではばたき飛行できないと思われますので,その半分としても12時間以上,おそらくその数倍はかかるでしょうし,さらに遠くへ飛ばないと別の陸地には着けません.したがって,エネルギー論的に見て,このような長距離移動ははばたき飛行だけでは不可能で,風の助けを借りないと実現できないでしょう.
以上から,日常の摂食飛行,パトロール飛行,産卵のための飛行など,敏捷に動き回る場合にはグルコースを使ったはばたき飛行を中心に行い,移住のような遠距離飛行の際には脂質を使った代謝に切り替えて,風に乗った滑翔を中心とした飛行を行っていると考えられます.ネットによく出ていますが,人間でも,マラソン選手は脂質代謝を行う方がより長距離をうまく走れると言われています.トンボも同じですね.
*1.アブのようにブーンという音を立てて非常に速くはばたく昆虫の筋肉の付き方は,トンボとは異なっています(間接飛翔筋の非同期筋).
*2.オーストラリアにのみ生息する大型のヤンマで,流水性.翅の前縁に褐色のラインがあることが特徴.

沖ノ鳥島近くに停泊する船で採集されたウスバキトンボ.
トンボの生態学 6.成虫の飛行移動
トンボの飛行による移動を分類する
トンボの移動手段は飛行です.落下など緊急事態以外歩くことはほとんどありません.飛行行動には,我々人間から見て,簡単にその目的を理解できるものとできないものがあります.もちろんトンボの方は,目的など意識できず,内因性の動機や欲求,刺激に対する反応としての行動をしているだけでしょう.しかし飛行の種類を定義し分類する際に,この認識の容易さを利用する方法があります.
まず第一は,例えば餌生物を見つけたので飛んで捕獲するとか,鳥に襲われそうになったので飛んで逃げるといったときの飛行です.こういった飛行は trivial flight と呼ばれています (Corbet, 1999).ここでいう trivial という語は通常「些細な」とか「つまらない」と訳されますが,数学では「自明な」という意味で使われます.後者の意味だとすると「見ただけでその目的や意味,実態が分かる証明不要な飛行」ということでしょう.このような明白な直接的目標と結びついた欲求行動(飛行)は,人の目の届く短時間で狭い範囲のものになるのが一般的です.おそらくそういう意味を含めて,trivial flight は「小規模飛行」と訳されています.
第二は,これ以外の,その目的や意味,実態が見ただけでは簡単には分からないような飛行です.こういった飛行は nontrivial flight と呼ばれています (Corbet, 1999).定義としては trivial flight 以外のすべての飛行ということになります.一般に長時間の広い範囲にわたる飛行は,人の目で全貌が観察できないということで,断片的な観察や知見を通して全体像を推察するしか方法がありません.おそらくそういう意味を含めて,nontrivial flight は「大規模飛行」と訳されています*1.
この大規模飛行については,さらに生息場所 habitat 内で行われている移動と,新しい生息場所への移動,つまり移住 migration を分けて考えます(Corbet, 1999).ただしこれらは概念的には区別できますが,実際は区別が難しくなる場合があります.この問題は,生息場所の範囲をどのように定義するかという点に集約されるように思います.そこで最初に,この問題に少し触れておきたいと思います.
トンボの生息場所は,生活史のステージによっていくつかの要素に分けて考えることができます.産卵が行われ卵や幼虫が生息する繁殖場所 reproductive site,成熟成虫が夜を過ごしたり,産卵意欲のないメスや未熟成虫が暮らすねぐら roost,採餌するための場所 foraging site,出会い場所 rendezvous,暑さなどを避けるための退避場所 refuge site などです.通常トンボは,一世代の間にこれらの場所間を移動しながら生活しています.したがってトンボの生息場所といえば,概念的にはこれらの集合体ということになります.
ここで,例えば池という繁殖場所について考えてみましょう.なわばりを形成するオスはそこに侵入した他のオスを追い払い,結果として一つの池に入れるオスの数に限りが出てきます.その結果その池に入れなかったオスが別の池に移動したとしましょう.そのときこの別の池は,この個体群の生息場所とみるべきか,この個体群にとっては「新しい」生息場所とみるべきかという問題が生じてきます.つまりこの移動は「移住」とみるべきなのだろうかという問題です.こういった問題は,生息場所の各要素のあらゆる場面で生じるでしょう.
Corbet (1999) はこの問題に対して次のような解答を示しています.彼は,上記の生息場所の各要素間の移動は,原則として,一世代内での「往復」または「往復の可能性が高い」移動であるという点に注目しました.対して移住は,原則として,一世代内での新しい生息場所への「片道」の移動であるということです.上記の問題は,この視点で答を出すべきだということになるでしょう.
一つの例を紹介します.山口県で行われたベッコウトンボの標識調査では,繁殖場所から半径1km以内では確認頻度が高いが,それ以遠では確認頻度が非常に低くなるという結果が出ています(三時・平田,1997).このことからベッコウトンボは比較的狭い範囲の生息場所を持つ,定住性が高いトンボだと言えるでしょう.兵庫県では,最後のものと思われる個体群が,2001年に,加西市の近接した2カ所に生息場所を持っていました.その各要素は,調査した限りでは,やはり池とその周辺数百メートルの範囲に限られていました.しかし同じ年,そこから約10km離れた小野市と加古川市でオスが1個体ずつ見つかりました(右欄外写真).周辺を調査しましたがあたりにはベッコウトンボの個体群は見いだせませんでしたので,おそらくこれらは加西市から飛来したものと思われます.これらは元の生息場所に帰るとは考えにくいので,定義によれば,これらは移住した個体と考えるべきでしょう.このように,この定義を厳密に運用すると,定住性の高いトンボにおいても,移住が起きている場合があります.
ベッコウトンボの例に限らず,いないはずの場所にある種のトンボが突然見つかるということはときどき見られます.一番はっきりとするのは,日本に分布していないトンボが見つかるという場合でしょう.Corbet (1999) はこれらはすべて移住した個体であるとし,多くの種の個体群は移動に関しては多型的(つまり一つの個体群の中に大なり小なり移住をする個体が混じっている)であることに留意すべきであると述べています.
では,以下で飛行の分類をし,その特性を見ていきたいと思います.
Ⅰ.小規模飛行 trivial flight
小規模飛行については,もうほとんど説明の必要はないでしょう.上に挙げた例以外では,生殖のための飛行があります.オオヤマトンボが池の周囲を飛んでパトロールする,エゾトンボのようなフライヤーがなわばりを巡回して飛ぶ,オオシオカラトンボのオスが産卵メスを警護するために飛ぶ,コシアキトンボがなわばりに侵入したオスを追飛するとか,活発な生殖活動にともなう飛行です(写真3,4).
*1.
Corbet (1962) は,トンボの生物学の以前の著作の中では,
tirivial の代わりに
appetential,
nontirivial の代わりに
non-appetential という語を使っています.
appetential には「欲求の」という意味があります.今回の
tirivial のほうが少し意味が広くなっているように思えます.

生息場所から10km離れた場所で見つかったベッコウトンボ.移住が確認できるトンボは,ベッコウトンボのように生息場所が限られる希少種であることが多い.
写真3.小規模飛行の例.左はオオヤマトンボの池周囲のパトロール,右はエゾトンボのなわばり飛行.こういった日の当たる場所をゆっくり飛ぶ場合,滑翔を多く用いているようだ.
写真4.小規模飛行の例.左はオオシオカラトンボのオスがメスの産卵を警護するためにメスを追って飛んでいる.右はコシアキトンボの追飛.このように激しい動きを伴う飛行は,ほとんどはばたき飛行である.
Ⅱ.大規模飛行 nontrivial flight (1) 生息場所内を移動する飛行
生息場所の内部で行われる大規模飛行には,処女飛行
maiden flight,通勤飛行
commuting flight,季節的退避
seasonal refuge のための飛行の3つがあります
(Corbet, 1999).
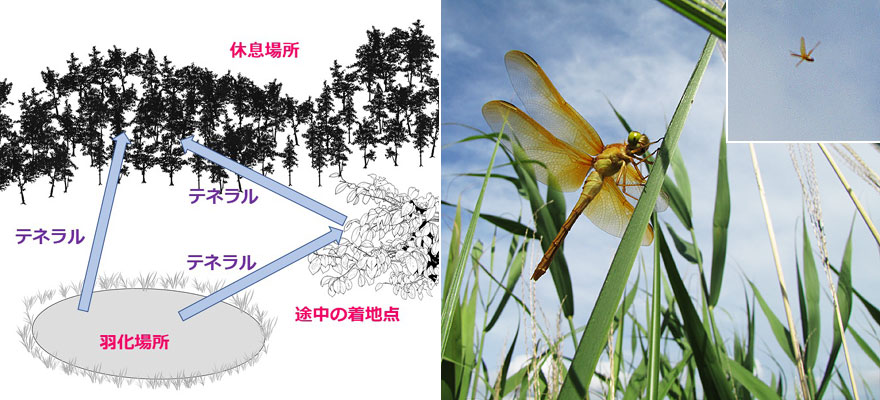
処女飛行は,羽化地点から休息場所までの飛行で,その距離は種によって大きく異なります.その距離が数十mを超えると,終着までを見届けることができませんので,終着点は推測になります(図2).
オオキトンボで,上空に舞い上がり,その姿が見えなくなるまで高いところへ飛んで行ったのを観察した経験があります(写真5).これなどはかなりの距離移動しているものと思われます.羽化殻から飛び立った後,このように途中で近くの葉などにいったん着地することがあります.こんな時でも,テネラルな状態で再び遠くに向かって飛び立った場合,休息場所,つまり成熟するまでを過ごす場所,に着くまでの飛行全体をさします.処女飛行は,一世代の間に1回だけ行われ,片道の,テネラルな状態での飛行であるという特徴を有します.
どのトンボでもほぼ例外なく,処女飛行は水面から遠ざかるような方向に飛びます.これが刺激に対する反応行動だとすると,水面は光を反射し水平な偏光をつくり出しますので,この偏光に対する負の走性のようなものがあるのかもしれません
(Corbet, 1999).
図2.処女飛行を示す模式図(左).写真5.オオキトンボの途中着地と処女飛行(右):遠距離を飛べるように体が硬化するのを待っているように見える.
オオキトンボは池で羽化した後に草むらに飛来し一時的に休止し,気温が高くなった頃に飛び立ち,目に見えなくなるまで上昇した.
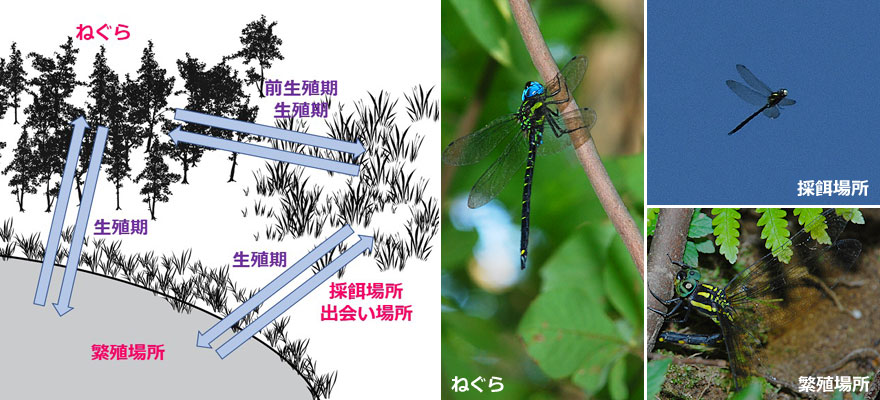
通勤飛行とは,ねぐら,繁殖場所,採餌場所,出会い場所の間の移動飛行のことです.これらの場所は,種によっては同一である場合があります.オスが水辺でなわばりを形成しメスを待つ多くのトンボでは,繁殖場所と出会い場所は同じになります.タイリクアキアカネではねぐらと出会い場所が同じです.ヒヌマイトトンボではこれら4つはすべて同じ場所のように見えます.その他いろいろな例があると思います.
通勤飛行は,前生殖期にある未熟成虫の場合や後生殖期にある老熟虫の場合は,主にねぐらと採餌場所の間の移動だけですが,生殖期にある成熟虫の場合,種ごとの生活様式によって,様々の飛行経路が存在すると思われます.各要素が離れている場合,その飛行全体を直接把握することはできません.ヤブヤンマの場合,実際に確認できるのは,ねぐらである暗い林内に休息している個体,摂食する個体が明け方や夕方に谷筋の摂食場所に現れしばらくすると山の方に飛び去ること,繁殖場所ではメスが林の方から飛んできて着地し産卵を始め終わったらまた林の方に帰って行くこと,オスは同じ場所で静止しメスを待っていることなどです(写真6).これらの断片的観察をつなげて,飛行経路を推測することになります.開けた場所などでは,通勤飛行そのものを観察することができる場合があります(写真7).
なお,処女飛行は,最初の通勤飛行ととらえることが可能です.
左:図3.通勤飛行の模式図.右:写真6.ヤブヤンマのねぐら,採餌場所,繁殖場所での観察.
写真7.左:アキアカネの繁殖場所へ向かう通勤飛行(往路).ねぐらと出会い場所が同じなので,タンデムではばたいたり滑翔したりして飛行する.
右:ウチワヤンマのメスの通勤飛行(復路).産卵を終えて一気に飛び上がりねぐらへ帰って行く.
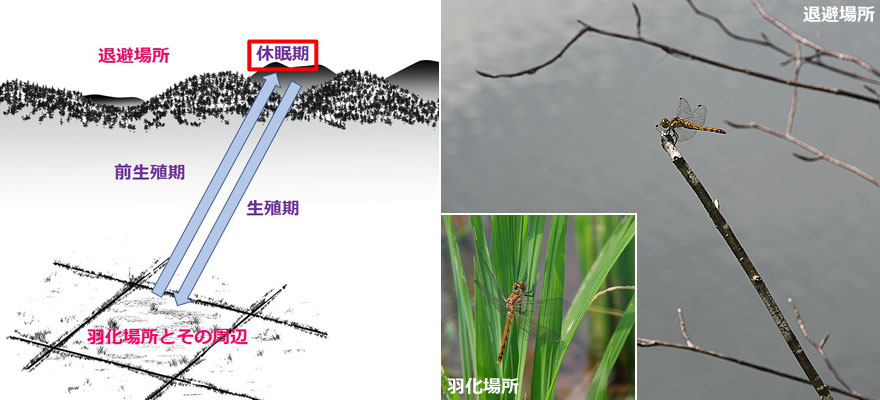
季節的退避とは,暑さ・寒さや乾燥を乗り切るための繁殖場所(羽化場所)から退避場所への移動,およびその逆の移動です.すべての種に見られるわけではありません.一般に,退避場所へ向かうときは前生殖期の未熟成虫で,退避場所から戻るときは生殖期の成熟成虫になります.季節的退避を行う種の特徴は,そのトンボにとって不適当な季節を「休眠」して過ごすという生活史を持っていることです.退避場所では,その大部分の時期を生殖休眠の状態で過ごしています.
日本のトンボには,乾燥を乗り切るための季節的退避を行う種はありません.これは主に雨季・乾季の存在する熱帯地方に生息するトンボに見られます.暑さを乗り切るための移動は,アカネ属,アオイトトンボ属,カトリヤンマなどに見られます.卵越冬をし,幼虫期間が短く,一年一化性のトンボばかりです.寒さを乗りきるために移動するトンボとしては,成虫越冬性の種が挙げられます.
アオイトトンボ属,カトリヤンマ,一部のアカネ属では,羽化場所から比較的近い林の中に退避します.この場合,処女飛行がそのまま季節的退避の飛行になっている可能性も考えられます.アキアカネやタイリクアカネなどは,よく知られているようにかなり離れた山間部に移動しますので,相応の長距離を飛行することになります.いずれも,その全貌が自明ではない nontrivial な飛行であるといえます.
図4.季節的退避の飛行の模式図.往路と復路の間に休眠期がはいるのが特徴.写真8.水田で羽化したアキアカネと高原の池で過ごすアキアカネ.
Ⅲ.大規模飛行 nontrivial flight (2) 移住-生息場所間の飛行
移住は,先に考察したように,どのトンボ種においても,個体群の一部に起きる可能性がありますが,移住が生活史の中に組み込まれていると考えられるトンボもいます.日本で見られるトンボではウスバキトンボがその代表例でしょう.もう例外なく毎年南方からやってきて日本で数世代を過ごしやがて死に絶える,というサイクルを繰り返しています.この「毎年決まったようにやって来る」というところに,移住することが生活史の一部として組み込まれていることが強く示唆されます.なおウスバキトンボの移住については,別の項でお話しすることにします.
そこで,移住は,そのトンボ種に生活史戦略として組み込まれている「絶対的移住
obligate migration」と,何らかの条件が整ったときに起きる「条件的移住
facultative migration」とに分けられています.前者は個体群すべての個体に環境条件に関わりなく必ず起きるのに対し,後者は,その条件が整った個体や時期にだけ起きることになります.
絶対的移住というリスクをともなう行動が進化した理由は,その方が生存の確率が高くなる(繁殖成功度が高くなる)ような環境に生息していたからと考えるべきでしょう.ではどのような場合に絶対的移住をした方が繁殖成功度が高くなるのでしょうか.一番に考えられることは,そのトンボにとって,生息場所がある時期に生存や繁殖に不適当になるような場合でしょう.しかしこれだけでは必ずしも移住が繁殖成功度を高めることになるとは限らないのです.
例えばアキアカネは,涼しい山へ季節的退避を行うトンボです.これは平地の夏の高温(生息環境として不適当)を避けるためというのが定説です.つまりわざわざ移住しなくても季節的退避によって不適当な環境となる時期(不適合期)を避けることができます.そしてまた環境が適合するようになったときに帰ってくればよいというわけです.ただし,不適合期はある程度継続するので,その間の生殖休眠が欠かせません.季節的退避の特徴に,退避中の休眠期の存在があるのはそのためです.
ただしここで注目すべきは,アキアカネが季節的退避によって不適合期を乗り切れるのは,厳格にコントロールされた「一年一化」の生活史を有しているからだということです.季節は通常一年周期で変化します.したがって,一年一化であれば一年周期の季節変化と同調させた生活史が構築できるというわけです.もう一つ,夏を越せるだけの高い山があるということも重要です.例えば,関西トンボ談話会で兵庫県淡路島のトンボ分布調査をしたときに,淡路島にアキアカネが極端に少ないことが指摘されました.これについて井上(1988)は,淡路島にはアキアカネが夏に移動できる適当な高山がないためと考察しています.
つまり季節的退避によって生息環境が不適合になる時期を乗り越えことができるのは,厳密にコントロールされた一年一化の生活史を有することと,往復移動できる範囲内に退避場所が存在することの,2つの条件が満たされたときであるということです.
以上から考えると,絶対的移住の方が有利となるトンボは,次のような条件の種だということになるでしょう.まず第一に,雨季と乾季,暑さと寒さなど,環境が適合する時期と不適合になる時期が存在し,往復移動できる近さに退避場所が
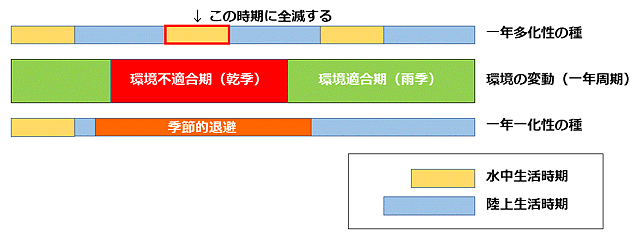
ない所に生息する種です.第二に,環境変動に生活史を同調させることが難しい化性を有すること,具体的には多化性ということになります(図5).多年一化性は一年中幼虫がいることになるので,こういう生息場所では生存できません.なお環境が適合している時期に幼虫期を過ごす必要があるので幼虫期が短いということも重要ですが,これは季節的退避でも同じです.
図5.一年周期の環境変化(雨季と乾季の例)と生活史の長さの関係.厳格にコントロールされた一年一化の種であれば,環境悪化の時期に季節的退避と休眠を行うことで,その場所で生活し続けることができる.しかし多化性の種はどこかで環境悪化の時期に幼虫期が重なってしまうので,別の生息場所に移住するしかない.
ウスバキトンボやヒメギンヤンマ
Anax ephippiger などの絶対的移住者は,このような条件を満たしています.
ところで,移住をするトンボたちはしばしば集合して大きな群れとなることが報告されています.例えばヨツボシトンボ原名亜種
Libellula quadrimaculata quadrimaculata の移住では,1個体が飛び立つと近くにいた他の個体に同じ行動が伝わるようすや,複数の小さな群れが次第に合流して大きくなっていくようすが観察されています.そして推定密度は400,000個体/km
2にもなったと報告されています(
Dumont & Hinnekint, 1973).
また,集団で移住飛行をしているトンボの群れには,別の種類のトンボや昆虫が混じっていることがあります.枝重夫博士が Corbet 博士とともに,カナダのエリー湖北岸の半島で南へ移住中(と彼らが推測している)のアメリカギンヤンマ
Anax junius の群れが休止している場面に遭遇しました.休止場所に同所的にいたトンボは20種類で,その中には
Tramea lacerata (ハネビロトンボ属)をはじめとした移住性のトンボが混じっていて,さらにサシバエやオオカバマダラなどの移住性の昆虫も群れで見つかっています(
Corbet & Eda, 1969).
最後に移住を行うことが知られているトンボを紹介しておきましょう.日本に飛来するものとしては,オオギンヤンマ*,アメリカギンヤンマ,ヒメギンヤンマ,ウスバキトンボ*,スナアカネ,ハネビロトンボ*,コモンヒメハネビロトンボ*などがあり,日本に定着しているトンボとしては,ギンヤンマ*,トビイロヤンマ*,アメイロトンボ*,ウミアカトンボ*,ベニヒメトンボ,ヒメトンボ*,ムツアカネ,エゾアカネ,さらに均翅亜目ではアオモンイトトンボ*やアジアイトトンボ*(世界的に見てもアオモンイトトンボ属のトンボに集中している)の名前も挙がっています(以上,
Corbet, 1999:Table 10.4, 10.5).

図5.日本で見られる移住を行うと報告のあるトンボたち(Corbet, 1999より).意外な種が混じっている.*印は南北大東島に分布.
沖縄から約400km離れた南北大東島は,その成立から一度も陸続きになったことがない海洋島です.島の基岩となる火山島は5,000万年以上前に形成されたそうですが,その後沈降し上部に珊瑚礁が形成され,100万年前ぐらいに今度は隆起を始めて現在の姿になった隆起珊瑚礁の島です(山口,2008).したがって原則としてそのファウナは移住によって形成されたはずです.そのせいでしょうか,南北大東島のトンボ相には上記の移住性トンボが多く含まれています(*印 津田,2000).そして,これ以外の南北大東島のトンボたちにも,ひょっとしたら移住する能力があるといえるかもしれません.例えば,近年分布拡大が見られる(た)リュウキュウベニイトトンボ,ムスジイトトンボ,アオビタイトンボ,コフキオオメトンボ(以上尾園ら,2012)などです.

南大東島を訪れたとき,こんなところにまでアオモンイトトンボはいるのかと驚いたが,移住するトンボならさもありなんという感じである.
1994年7月 南大東島在所.
トンボの生態学 6.成虫の飛行移動
トンボの飛行を助ける気流
ここでは,トンボの飛行を助ける,気流(風といってもよい)について考えてみたいと思います.はばたき飛行はエネルギーを多く使う飛行で,長時間続けられないことはお話ししました.大規模飛行に限らず,小規模飛行においても,ヤンマ科,オニヤンマ科,ヤマトンボ科などはかなり長い時間飛び続けています.エネルギー消費という観点で,飛行全般において,滑翔による飛行エネルギーの節約には重要な意味があります.実際トンボが飛ぶのを観察していると,はばたかずにグライダーのように滑翔する姿がよく見られます.ちなみにグライダーは,上昇気流によって高度をかせぎ,その位置エネルギーを運動エネルギーに変えながら飛ぶそうです.もちろんそのままではついには地上に降りてしまいますから,再び上昇気流をとらえて位置エネルギーをかせぎ,これを繰り返すのが長い距離を飛ぶ秘訣だそうです.
おそらくトンボの場合も,同じような原理を使っているのだと思います.ただトンボは,はばたき飛行という動力飛行もできるので,気流の状態が好ましくなくても飛び続けることが可能だといえます.実際は状況に応じて,はばたき飛行を適当に使いながら,滑翔を中心として飛んでいるのでしょう.秋の午前,アキアカネの通勤飛行をみていると,タンデムペアが高いところを滑翔しているかと思うと,風が吹いたり方向転換するときなどにはばたき飛行を行って航路を調整しながら,産卵場所に向かって飛んでいくのが観察できます(写真8).またコシアキトンボの摂食飛翔のような狭い空間を飛ぶ場合でも,滑翔を多く利用して飛んでいます(写真9).
写真8.アキアカネの通勤飛行を連続撮影したものを使って合成.G:滑翔,F:はばたき飛行.2018年10月18日 9:46.
写真9.コシアキトンボの摂食飛翔を連続撮影したものを使って合成.G:滑翔,F:はばたき飛行.2018年6月1日 15:34. 滑翔中にローリング(左右の揺れ)が起きているのが分かる.これは気流の小さな乱れによる.
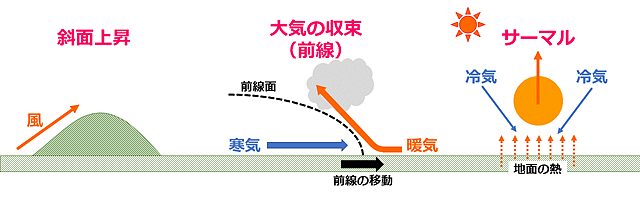
さて,滑翔を助ける上昇気流はどのようなところで発生するのでしょうか.よく知られている3つの例をおさえておきましょう.一つは風が山などの斜面を上っていくときに生じる斜面上昇,二つ目は大気の収束(前線)による上昇,三つ目はサーマルの上昇です.
図6.上昇気流を生じさせる3つの主な要因.
このうちトンボの長距離移動にとって重要な役割を果たすのは,大気の収束とサーマルです.斜面上昇というのは昆虫を高い山に吹き上げる効果はありますが,長距離移動の出発のための上昇気流となることはないようです (
Corbet, 1999).
一方サーマルは,小規模移動から大規模移動に至るまで,トンボの滑翔に重要な役割を果たしているように思えます.サーマルは,日射によって地面のある部分がその周囲より強く熱せられ,その付近の空気が温められて生じる空気塊です
*1.周囲より温度の高い空気塊(サーマル)ができると膨張し密度が小さくなって,大気圧により浮力が働き上昇を始めます.太陽高度が高くなり日差しが強くなるほど,力強い上昇気流を生み出します.トンボが晴れた日によく飛ぶのは,体温が上がる以外に,サーマルの発達と関係があるのかもしれません.
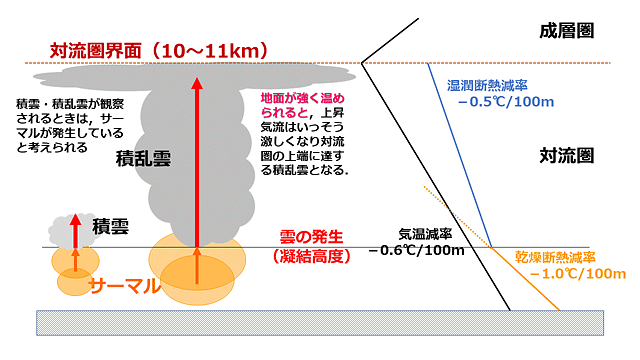
風がないとき,サーマルは断熱膨張しながら上昇していきます.このとき100m上昇するたびに約1℃温度が下がります(乾燥断熱減率).一方気温は一般に高いところほど低くなりますが,こちらは100m上昇するたびに約0.6℃温度が下がります(気温減率).サーマルが上昇し始めたときはまわりより温度が高いのですが,サーマルの方が気温の下がりかたが大きいので,(雲が発生しない限りは)やがてまわりの気温と同じになるときがきます.こうなるとサーマルはもはや上昇できなくなります.
一方雲ができるときは少し状況が違ってきます.サーマルが上昇すると,ある高さ(凝結高度)で雲が発生する場合があります.雲が発生してもサーマルは上昇し続けますが,水蒸気が凝結して雲になるときに凝結熱(潜熱)が放出されますので,その分気温の下がりかたが鈍くなります.雲ができながら上昇する場合,100m上昇するたびに約0.5℃気温が下がります(湿潤断熱減率).したがって気温減率より小さくなって,原理的にはサーマルはどんどん上昇し続けます.夏の暑い日など地面が強く熱せられ上昇気流の強いサーマルが発生した場合,対流圏界面まで上昇しながら雲を発達させます.これが積乱雲です.そこまで強いサーマルではない場合は積雲をつくります.積雲と積乱雲は対流雲といってサーマルによってつくられる典型的な雲です(図7).ちなみにグライダーを飛ばすときは,積雲が出ている日がいいそうです.
図7.雲ができる場合のサーマルによる上昇気流.気温減率,湿潤断熱減率,乾燥断熱減率などの数値は条件によって変化するので,あくまで原理的な理解の図だと思ってほしい.
写真10,11.積雲と積乱雲.上が積雲で雲底が一定の高さにあるのが分かるだろう.この高さが凝結高度.下の積乱雲は金敷雲とも言われ,上部が対流圏界面に一致し平らになる.
サーマルは,トンボだけでなく,チョウやバッタなど,渡りをする昆虫を上昇させるのにも役立っています.例えばオオカバマダラという有名な渡りをするチョウが2,300mまで持ち上げられた記録があります(
Strauss, 1985).トンボでも,アメリカギンヤンマが高度2,300mの高さのところを力強く飛んでいた記録(
Glick, 1939)や,4,000mの標高にある氷河にトラップされていたアメリカギンヤンマの記録(
Papp, 1974)なども,この上昇気流で説明が可能です.
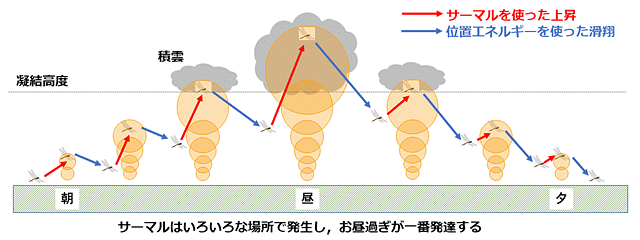
サーマルはトンボを上昇させるだけでなく,トンボはこれでかせいだ位置エネルギーを使って降下しながら滑空前進できます.そして次のサーマルをとらえることができたらまた上昇して同じことを繰り返せます.うまくサーマルをとらえられなければ,はばたき飛行をしてサーマルを探せばいいでしょう.こうやって継続的省エネ飛行が可能になります.また,サーマルは午前中は太陽高度の上昇とともに発達してそのてっぺんが高くなっていき,午後は逆に低くなっていきます.つまり次々とサーマルにうまく乗れば,高さも自動的に調整されることになります(図8).
図8.よく晴れた風のない日に,サーマルだけを使って省エネ飛行(滑翔を中心とした飛行)する場合の理論図.あくまで理屈だけで考えたもので,実際にこのようになっているかどうかは分からない.横軸は時刻とともに位置の変化も表している.サーマルの発達が時刻によって異なるので,サーマルを渡り歩いていけば,夕方には地上付近に到達する.図では雲の中を飛行しているように描いているが,雲の下を飛んでいるかもしれない.必要に応じてはばたき飛行も利用することになる.Corbet (1999) の記述を参照して描画.
実際さまざまの飛行で,サーマルをどれほど利用しているかはトンボに聞くしかありません.サーマルを利用しなくても,はばたき飛行で高度をかせぎ,その位置エネルギーで滑翔することも可能だからです.おそらくトンボは,サーマルを利用できるときは利用し,そうでないときははばたき飛行をして上昇し,滑翔しているのではないでしょうか.
さて,次にトンボの飛行にとってもう一つの重要な気流をつくりだす,大気の収束についてみていくことにしましょう.大気の収束帯,つまり前線は,異なる性質を持つ空気がぶつかったところにできます.また前線ができるところは,前線に向かって風(収束風)が吹くので,移動昆虫に水平方向の駆動力を与えます.小規模飛行の場合や,大規模飛行でも,処女飛行や通勤飛行などでは,サーマルだけでも十分距離が稼げそうです.しかし,海を渡るような長距離飛行の場合は,水平方向の駆動力の助けを借りないと,速度が出ないと思われます.
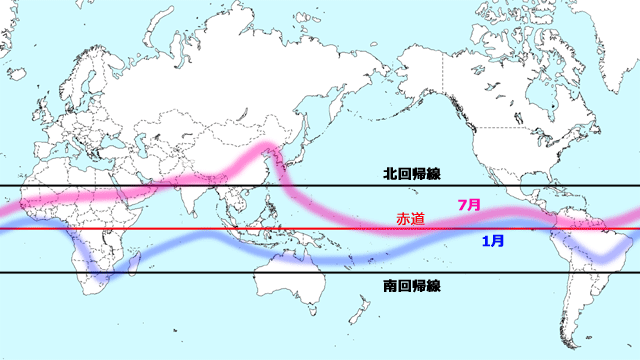
日本では,収束帯は,温暖前線,寒冷前線,停滞前線などの形でみられます.温帯地域の収束帯は不安定で生成消滅を繰り返します.しかし地球規模で安定した収束帯がみられる地域があります.それは熱帯です.熱帯収束帯(ITCZ)と呼ばれるこの巨大な収束帯は,基本的に北東貿易風と南東貿易風がぶつかったところにできますが,その位置はモンスーンの影響も受けています.熱帯収束帯では激しい上昇気流が生じ,雲が発達してたくさんの雨が降ります.水を必要とするトンボやその他の昆虫にとっては非常に重要な意味を持つ場所です.また熱帯収束帯は太陽のみかけの動きにともなって,夏は北の方に,冬は南の方に移動します(図9).
図9.熱帯収束帯の大まかな位置とその移動.さまざまな資料をもとに作成.アフリカ東部から太平洋東岸にかけて,1月と7月の位置が大きく開いている.これはこの地域に吹くアジアモンスーンの影響を受けているためと考えられる.つまり北半球の夏にはアラビア海やインド洋から大陸に向かって,同じく冬には大陸から海に向かってモンスーンが吹くので,熱帯収束帯がそれぞれ北や南へ押されていると考えられる.7月には,この東端に位置する日本付近まで熱帯収束帯がやってきていることに,南方からの飛来種という観点で注目したい.
熱帯収束帯の移動とともに移動する昆虫にサバクトビバッタがいます.バッタは農作物に甚大な被害を及ぼしますので,その飛来が古来から記録され,最近もよく研究されています.アフリカ北部では,春から夏にかけて,熱帯収束帯の北上とともに,南からの収束風に乗ってサバクトビバッタの群れが北上し,秋になると今度は熱帯収束帯の南下にともなって,北からの収束風に乗ってサバクトビバッタも南下し,アフリカ東部に移動するそうです(Williams, 1958).
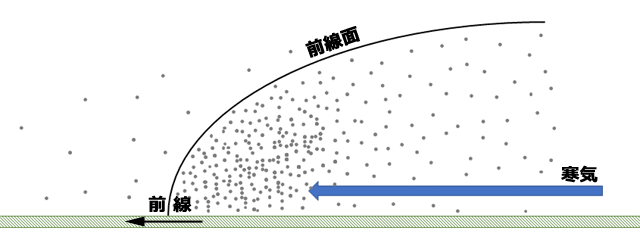
収束帯は,飛行している昆虫に,収束風によって水平の駆動力を与えるだけでなく,その昆虫を集合させる効果があるといわれています.イギリスの
Schaefer (1976) は,寒冷前線の通過にともなう飛翔昆虫の密度を,レーダーを使ってとらえています.それによると,寒冷前線の通過後,通過前より飛翔昆虫の密度が60倍にも増加したそうです.つまり寒冷前線に向かって吹く寒気が,飛翔昆虫を前線面の下側に集結させているのです.
図10.寒冷前線によって飛翔昆虫が集合してくるようす.Schaefer (1976) を参考にして描画.
さて,主に上昇気流について述べてきましたが,水平方向に吹く風がトンボをはじめとした昆虫の移動にとって重要なことは言うまでもありません.気圧傾度によって吹く風,モンスーン(季節風),海風や陸風など,これらの風が追い風となって,昆虫に水平方向の駆動力を与えています.日本にやって来る台風は熱帯収束帯で発生します.したがって,熱帯収束帯に集まってきたトンボは,台風によって運ばれる可能性があります.
より高高度を吹く,地衡風,偏西風,ジェット気流などが昆虫を運ぶかどうかは,なかなかその証拠を直接つかむことができませんが,ネパールのクーンブ・ヒマール山群というエベレストの近くで,標高6,300mを飛ぶウスバキトンボが目撃されていて,さらに恒雪帯と呼ばれる氷河が発達しているような高度まで移住飛行の記録があるということです(Wojtusiak, 1974).
*1.サーマルは周囲との温度差ができるようなところによく発生します.林縁や水辺などに開放的な地面があるような場合です.その大きさは,温められる地面の大きさに影響され,さまざまです.一方海のように一様に温められる場所では発生しにくくなります.サーマルの上昇にともなって,その周囲には下降気流ができます.
トンボの生態学 6.成虫の飛行移動
ウスバキトンボの話
さて,理論的な話ばかりが続きました.最後に,実際のトンボの移住についてお話ししてみたいと思います.日本に分布するトンボの中では,なんといってもウスバキトンボが絶対的移住者としては有名です.
ウスバキトンボは,よくコスモポリタン種(世界を股にかけている種)であると言われてきました.最近の研究で,世界中のウスバキトンボに遺伝的交流がある,つまりわかりやすくいえば,世界中のウスバキトンボが一つの繁殖集団を形成している可能性が示されています (Ware et al., 2022).一般に広域分布種は各国や地方で個体群が生殖的に隔離された状態にあることが多く,それぞれの個体群で独自の進化
*2が起きるために,個体群ごとに特徴的なDNAの塩基配列(ハプロタイプ)が見られることが多いのです.しかしウスバキトンボには,地域独特に発達したハプロタイプがあるようには見えないようです.これは,どこかで生じた突然変異が,移住と自由な交配を通じて,世界中の個体群に広がっていくため,と考えられています.
こんな世界を股にかけているウスバキトンボですが,兵庫県では,春になると南方から飛来しはじめ,初夏,そして秋口に大発生します.全国的には,暖かい期間中にはどんどん北の方に分布が広がっていき,北海道の最北部にまで到達します.さらに北へも飛んでいっているようですが,詳細は分かりません.しかし冬の到来とともにこれらはすべて死滅すると考えられています.幼虫が越冬できるのは,日本では八重山諸島ぐらいのようです(尾園ら,2012).
卵期は5日,幼虫期は34日(関西トンボ談話会,1984)という記録があり,産卵から次の世代が誕生するまで,水中生活期間がわずか1ヶ月少々という短期間です.日本の気候下では,地域によって,2~5世代ぐらい繰り返せる計算になります.この水中生活期間の短さや多化性は,すでに述べたように,絶対的移住者の必要条件です.また翅の肛角部が大きく後方に広がっているのは,滑翔のための適応的形質であると考えられています(
Corbet, 1999)(右欄外図).
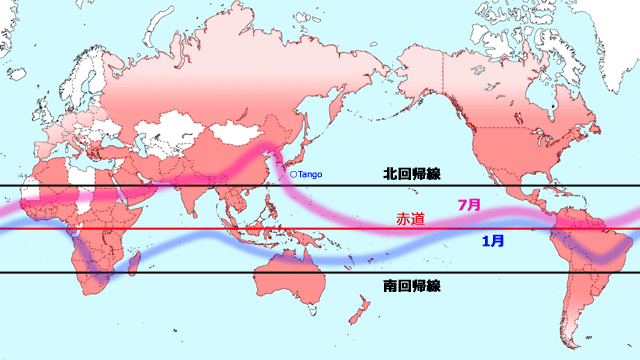
日本において,ウスバキトンボが太平洋上を飛んでいるというのは,過去にもときどき海上での発見によって報告されていました(例えば,野平,1960).しかしこれを初めて計画的に行ったのは,南方定点の定点観測船上での調査です(朝比奈・鶴岡,1967, 1968, 1969, 1970;橋本・朝比奈,1969).南方定点は,北緯29度,東経135度にあり,
Tango と呼ばれています(図11).潮岬の南方約500km,最も近い陸地である足摺岬・種子島から450kmほど離れた太平洋上です.観測船は二交代で観測に当たり,流されても最大直径約90kmの範囲内にとどまるように決められています.以下,これらの文献から引用しながらお話を進めます.
図11.ウスバキトンボの分布・飛来記録.図9に津田(2000)に挙げられている国々を塗りつぶして作成.一国に一つでも記録があれば国全体を塗りつぶしているので,記録の非常に少ない高緯度地域(ヨーロッパ,ロシア,カナダなど)は淡色で表示した.全体として,熱帯収束帯とその周辺が分布中心になっていることが分かる.
フランス・スペイン以外のヨーロッパの国々は,Web サイト DragonflyPix.com (2022.4.21現在) を参考にして追加,また津田(2000)には韓国が欠落していたので,李(2001)で確認して追加した.熱帯の一部空白部分は津田(2000)には国名が挙がってなかったが,おそらく分布していると見てよいであろう.
Ⅰ.日本の南方定点 Tango における調査
この調査は,1966年から1968年までの3年間行われ,全部で5種のトンボが確認または採集されました.ウスバキトンボ,ハネビロトンボ,ギンヤンマ,オオギンヤンマ,アジアイトトンボです.ウスバキトンボがほとんどを占めています.ギンヤンマは20頭以上確認されており,海上をかなり移動していることが判明しました.オオギンヤンマは2頭,ハネビロトンボは30頭以上確認されています.またアジアイトトンボが4頭とれていますが,これはセジロウンカやトビイロウンカというイネの害虫と一緒になって飛来していました.
面白いことに,夜間,灯火に飛び込んでくる個体が結構たくさんいます.1968年8月1日の調査ではウスバキトンボ48頭,ハネビロトンボ8頭,ギンヤンマ2頭がライトトラップで採集されました.しかもこれらのトンボは,すべて夜半過ぎ,月が沈んだ後に灯火にやってきました.このときの月は上弦を過ぎ満月に至る途中で,日没後から月光が海面を照らしている状況でした.トンボは月光を感じているのかもしれません.それにしても夜間も移住飛行をしているというのは驚きです.
風速は飛来数にあまり関係がなく,風向については西南の風に支配されていて,飛ぶ向きは風の向きに一致していると書かれています.ただデータからは,特に秋になると北から東寄りの風で飛来する場合が若干多くなるように見えます.しかもこの時期に一緒に飛来するトンボ以外の昆虫には,夏季の南方種と違って西南日本に見られる普通種が多いと書かれていて,ひょっとすると秋には北から南への移動が起きているのかもしれません.
風に乗ってきたウスバキトンボは,ときどき船に止まったり,風の弱い日は船体の風下側を巡回飛行する性質があるようです.またウスバキトンボが飛ぶときは,積雲や積乱雲といった対流雲が出ているときと書かれており,上で述べたように上昇気流が生じているときであることが分かります.やはり省エネルギーの滑翔を中心とした飛行を行っているのでしょう.
この調査では全部で90種類ほどの昆虫が確認されていて,その中にはウンカなどの餌となる小昆虫もたくさん含まれています.しかしトンボを捕まえて解剖してみても,素嚢が空で,飛行中に摂食している確証は得られていません.飛来した多くのトンボは成熟していて,捕まえて三角紙に入れると放卵するメスもいましたが,飛行中に交尾態になっている個体は観察できていません.
ウスバキトンボに関しては,面白い現象が報告されています.一つは1967年7月24日のことです.12:20から30分間ほど,
台風10号の目が観測船の真上に位置しました.このとき,
上空が真っ黒になるほどたくさんのウスバキトンボが飛んでいたというのです.台風がトンボを運ぶことを初めて目視確認できた瞬間です.二つ目は,1968年9月17日,この日は非常に多くのウスバキトンボが飛来しました.ライトトラップでも215頭が採集されています.この日,にわかに信じがたいことですが,
ウスバキトンボが海上に静止していて,船が流されるのにしたがって飛来した,と書かれているのです.
観測船は結構大きく流されて移動しているのですが,どの地点においても飛来にあまり差が感じられなかったということなので,ウスバキトンボは,平面的にはかなり広い範囲を飛んでいるものと推察されます.特に船を目指しているとは考えられないので,この海域を飛んでいるウスバキトンボは相当の数になると考えられます.
以上のように,この調査から得られることは非常に興味深く,情報は多いのですが,目の届く範囲の観察にとどまっているので,高高度を飛ぶ可能性については述べられていませんし,ウスバキトンボの出発地については,風向きや気圧配置から推定するぐらいしか方法がありません.ところが近年,ウスバキトンボが1,000m程度の高高度を飛んで移住する可能性と,その出発地をかなりはっきりと特定した研究が発表されました.次はこれらの研究を紹介したいと思います.
Ⅱ.西インド洋におけるウスバキトンボの移住の研究
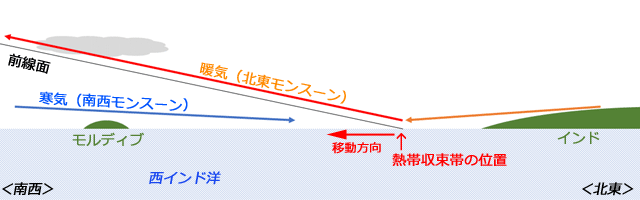
これらの研究は,インド洋に浮かぶ1,200もの珊瑚礁からなる国,モルディブ共和国で行われました.国のほとんどが珊瑚礁ですから,モルディブにはトンボの生息に適した淡水域はほとんどありません.そういう意味で,モルディブでの観察は,洋上の船上での観察に近いものがあります.そのモルディブでは,昔から,10月のはじめになると突然トンボの大群が現れることが知られていました.そしてこの10月のトンボの大群の出現が,まもなく北東モンスーンが吹き始める予兆として,地元の人たちに知れ渡っていました.
モルディブが存在する西インド洋はモンスーン気候で,5月から9月には南西モンスーン,11月から3月には北東モンスーンが吹きます.10月というのは,ちょうどその端境期に当たり,モンスーンの向きが変わる季節なのです.そしてそのころ熱帯収束帯がモルディブを通り過ぎて南へ移動します.同様に半年後の4月もモンスーンの端境期で,今度は熱帯収束帯はモルディブを北へ移動していくことになります.
まず
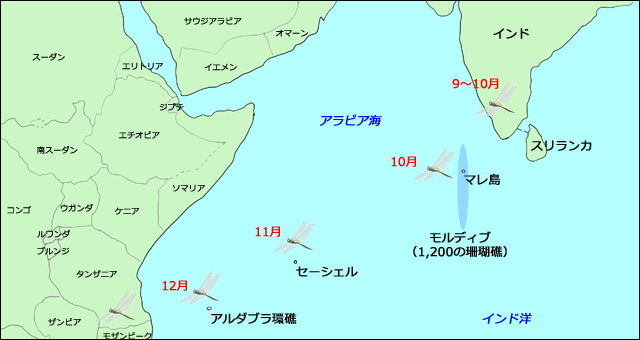
Anderson (2009) は,過去のさまざまの知見,それまでの自身の観察,地元の観察者とのやりとりなどから,ウスバキトンボは,インド南部で9から10月にかけて大量に南下を始めること,そしてモルディブに10月に最初に到着し(インドから600~1,000km),セーシェルに11月(インドから2,700km),アフリカのアルダブラに12月(インドから3,800km)に出現するという知見をまとめて,ウスバキトンボが,インドから西インド洋を渡って,アフリカに移動するという仮説を立てました.さらにこれが熱帯収束帯の南への移動と軌を一にしていることから,南下する熱帯収束帯の背後に吹く北西モンスーンが,ウスバキトンボを運んでいると考えました.
図12.西インド洋,アラビア海付近の地図.モルディブは1,200もの南北に長く連なる珊瑚礁からなり,アルダブラは環礁の島である.セーシェルはマエー(Mahe)島が首都のある一番の大きな島で,115の珊瑚礁と小さな島々からなる.
その移動の中で,特にモルディブにウスバキトンボが飛来するタイミングに彼は注目しました.モルディブに最初にウスバキトンボが現れ,さらに数が増えていく10月上旬,毎年のようにモルディブではまだ南西モンスーンが吹いています.つまりトンボの移動方向と逆なのです.600km以上もあるこの海洋上を逆風で飛んできているのか? そこで彼は,もっと上空を飛んできているのではないかと考え,情報を収集しました.するといくつかの面白い情報や現象を見つけました.
熟練した気象観測者であるモルディブ気象局長からの連絡で,マレ島に隣接するビリンギリで,多数のウスバキトンボが,2008年の10月後半の16:00ごろ,大量に上空から降りてくるのを観察したというのです.局長は,双眼鏡で垂直に上向きに観察したところ,降りてくるウスバキトンボを1,000mの高さまで確認することができた(つまりそれ以上の高さから降りてきている)ことを
Anderson に告げています.
また
Anderson は,上空を流れる雲の動きに注目しました.2003年,10月4日には,高高度の雲も低高度の雲も南西の風に流されて北東の方に動いていました.ところが10月5日になると,低高度の雲はまだ北東に流れていましたが,高高度の雲は逆の南西に流れていました.そしてその日にこの年最初のウスバキトンボがモルディブに出現しました.つまり高高度のこの風に乗れば,南西モンスーンが吹いているときでも,インドからモルディブにやって来ることができるわけです.
そこで彼は,マレ島の隣のフレル島で日本の海洋研究開発機構が2006年に上げたラジオゾンデのデータを収集して確認してみました.データは10月18日(この年ウスバキトンボが初めてマレ島に飛来した日)から11月26日までのものでした.これによると,高度1,200~2,400mに3~4m/sの東北東の風が吹く層と,2,900~5,000mに5~6m/sのもう一つの東北東の風が吹く層があることが分かりました.やはり高高度に逆向きの風が吹いていたのです.
これについて,彼は次の図ように考察しています.10月は,南下する熱帯収束帯がモルディブを通過する時期です.この時期まだ熱帯収束帯の北側の気温が高いので,ちょうど温暖前線が進むような感じで南に移動していきます.熱帯収束帯(前線)がモルディブより北に位置しているときには,その前線面はモルディブの上空にあって,そこには前線面を吹き上がる暖気が北東風となって吹いていることになります.これが高高度の風で,低高度の風はそれに対抗して吹いている南西モンスーンです(図13).
図13.熱帯収束帯がインドとモルディブの間を南下するときには,上空には北東の風が吹き海面付近は南西の風が吹く.
ラジオゾンデによって測定された風速をもとに,仮に5m/sの風が吹いていて,その風とともにウスバキトンボが移動しているとするならば,計算上一日で432km移動することになり,インドとモルディブマレ島の距離を600kmとした場合,1.39日でその距離を移動することになります.驚くべき速さです.熱帯収束帯はこの間を移動するのに約8日かかるそうですので,熱帯収束帯が通り過ぎる前にウスバキトンボがやって来ることに不思議はありません.地元の人が言う「ウスバキトンボの大群がやって来ると北西モンスーンがまもなく吹き始める」というのも理解できます.
ウスバキトンボが高高度を飛ぶというのは,中国の研究者
Feng (2006) も述べています.夏に中国の渤海湾
Bohai Gulf を渡るウスバキトンボは,夜間に,1,000mの高度に至るまで飛んでいて,一番密度が高いのは温度逆転層のある200-300m,および500m付近であったと,レーダーを使った観測で確認しました.また彼は,ウスバキトンボの動きから,一日に 150-400km 移動するとも述べています.
ところで,ウスバキトンボが高高度を飛ぶのはほぼ確実なようですが,このモルディブのウスバキトンボの出発地がインド南西部であるという彼の予測は,やはりまだ予測のままです.この問題に関して
Hobson et al. (2012) (この中には
Anderson も含まれています)は,安定同位体分析法という技術を使って,このウスバキトンボの出発地をかなり正確に見いだすことができたと報告しました.
彼らは安定同位体として重水素(
2H)を用いました.重水素は,水分子(H
2O)の水素原子として,自然界の表層水や雨水の中に一定の割合で含まれています.ただし,地域によってその割合が微妙に異なっているのです.まず植物がその水を使って光合成を行い,やがて食物連鎖を通じてトンボの体内に入って代謝され,体を作る物質中に蓄積されていきます.したがって,体の構造をつくる物質中の水素原子には,幼虫の育った地域の重水素の割合が反映されることになります.彼らは,成虫の翅を使って,重水素の割合を分析をしました.標本の採集はモルディブのマレ島とクンフナッドホー環礁で行い,雨水や表層水はインド亜大陸の各地のものを分析しました.
その結果モルディブのウスバキトンボの起源は,
Anderson のインド南部という予測と少し異なって,インド北部,ネパール,さらにより遠くのアジア地域であると推測されました.つまりこのウスバキトンボは,インド亜大陸横断という,より長距離の移動を行って,モルディブにやってきているということになります.
Ⅲ.熱帯収束帯の移動と連動するウスバキトンボの移動
Anderson の指摘している,ウスバキトンボの熱帯収束帯に関連した移動については,旧くから指摘されてきたことです.例えば,
Corbet (1962) は彼自身の観察として,「赤道直下のウガンダのエンデベに(ウスバキトンボが)数多く現れるのは3,4月と9,10月の年に2回,南緯3度40分のタンザニアのシンヤンガでは12月と1月に最も多くなる.これらの時期は,これらの地域における熱帯収束帯の通過時期に一致する.またモザンビークのベイラで,(熱帯収束帯が通過する時期の)雨が降っている1月中頃に,ウスバキトンボが飛んでいくのを見ている」と述べています.
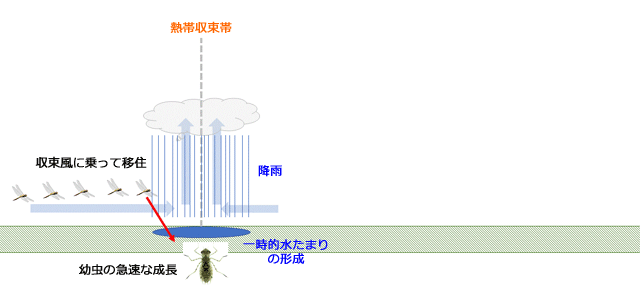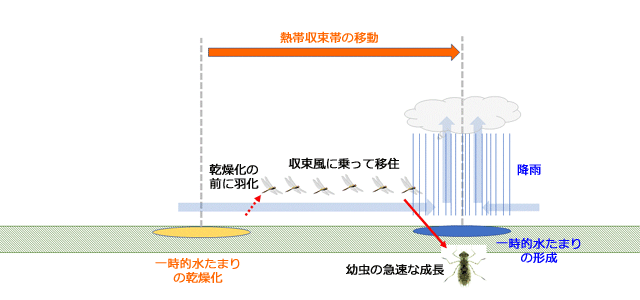
では,熱帯収束帯とともに移動していくことに,どのような意味があるのでしょうか.Corbet (1962,1999)は,熱帯収束帯は上昇気流が生じ降雨が多いところであるので,熱帯収束帯と一緒に移動すれば,水のある場所に自動的にたどり着くことができる,という適応的価値を強調しています(図14).その結果,雨季や乾季が存在する地域において,風に乗って絶対的移住をするという行動形質が進化したと考えています.彼はそのことを示唆する観察として,ウガンダで,羽化した場所がまだウスバキトンボが集まって繁殖活動するのに敵した状態であるにもかかわらず,羽化と同時に姿を消してしまったという観察例を挙げています.
図14.熱帯収束帯に吹き込む収束風に乗って移動すれば,自動的に水のある場所にたどり着けることを示すアニメーション.降雨によってできるのは一時的水たまりですから,卵期や幼虫期の発育速度が高く,水たまりが干上がる前に羽化する必要がある.
熱帯収束帯は季節によって南北へ移動します.したがって,熱帯収束帯とともに移動するウスバキトンボも,南北への往復移住を行うことになります.実際,上記の
Corbet (1962) の観察はそれを示唆しています.先の
Anderson (2009) も,いくつかの状況証拠からアフリカからインドへの復路移住の存在を示唆し,往復移住の可能性に言及しています.
以上から,ウスバキトンボは,熱帯収束帯の収束風に乗って移動することが繁殖成功度を高めるという選択圧が働いた結果,羽化するとまもなく風に乗って移動するという行動形質が進化し,絶対的移住者となったと考えることができるでしょう.熱帯収束帯にともなって南北に移動するという形質は,熱帯収束帯の移動地域内で生活する限りにおいて非常に有効であると思われます.
Ⅳ.日本にやって来るウスバキトンボの意味するもの
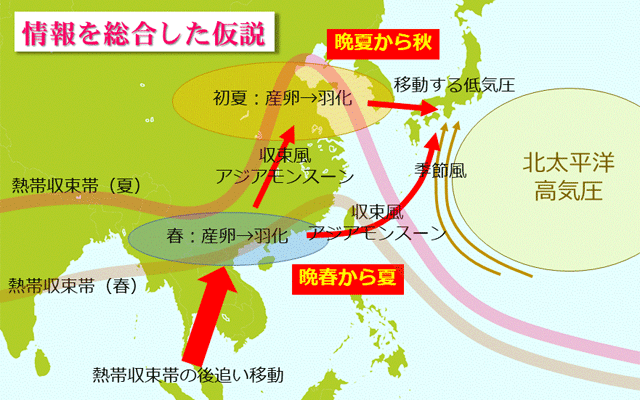
日本にやってくるウスバキトンボはどこからやってくるのでしょうか.これについては,日本のウスバキトンボを使った安定同位体分析が行われています
(Hobson et al., 2021).日本で採取されたサンプルは,鹿児島,滋賀,静岡,埼玉,宮城,北海道の6カ所で,安定同位体としては,翅に含まれる重水素を用いています.それによると,4月ごろ鹿児島にやってきている個体は,ミャンマー北部から中国南部,ボルネオのスラゥエシあたりからやってきているらしく,夏になるとインド北部やチベット高原西側あたりから飛来するようになるようです.さらに夏の終わりから秋になると,中国北部や朝鮮半島などの広い地域からやって来ている可能性が高くなるということです.
図15.Hobson et al., (2021) を中心に,今までの情報をまとめて,日本にやってくるウスバキトンボのルート(赤矢印)を描いてみたもの.あくまで一つの仮説である.
春から夏にかけての飛来元となる地域は,図11を見て分かるように,いずれも熱帯収束帯の移動範囲内です.シーズン終わり近い夏から秋にかけての飛来元である中国北部や朝鮮半島は,このあたりに南方から飛来したウスバキトンボの次以降の世代が起源となっているのでしょう.
また
Hobson et al. (2021) によると,よく言われているように,日本に飛来したウスバキトンボが国内で繁殖・羽化し,その子孫が順次北上していくということではないようです.特にシーズン前半ではほとんどが海外から移住してきた個体であり,後半になっても,国内で羽化した個体の割合は,海外から移住してきた個体より低いということが分かっています.これの意味することは,日本で羽化した個体たちは,国内を順次北上するというより,海外へ移住してしまうものが多いということでしょう.つまり日本のウスバキトンボは,日本で羽化しても海外へ行ってしまい,それを補充するのは海外からの移住者だということです.これが事実なら,日本でマーキング調査をしても,国内で再捕獲される可能性は低いということになりますね.
さて,起源がどこであれ,熱帯収束帯が北に位置するような場所でウスバキトンボが繁殖したとします.その個体群が羽化する時期に,たまたま北向きに風が吹いていたとしたら,このウスバキトンボたちは,その先天的な行動形質にしたがって,熱帯収束帯に向かわず,逆の北方向に飛んでいくことになるはずです.
夏には熱帯収束帯は日本のすぐ南にまで北上してきています(図11)から,こういった事態は,小笠原高気圧が発達し,それを回り込むようにして南寄りの季節風が吹く日本付近では特にありそうです.これがウスバキトンボが毎年のように日本に飛来する原因ではないでしょうか.現に,南方定点
Tango (ここは熱帯収束帯の位置より北になります;図11)での観察で,5月から8月ぐらいまで,南西方向から吹く風に乗って多くのウスバキトンボが北東の方に飛び去っています.
日本付近では,この南からの季節風が北の寒気と衝突し,梅雨前線を形成して,その一帯では雨が多く降ります.そこでは一時的にウスバキトンボの戦略が成功しています.その後梅雨前線は次第に北上し,日本は夏を迎えます.これが,ウスバキトンボの飛来前線が日本列島を北上することにつながっているのでしょう.ちょうど前線が生成消滅する一帯が,熱帯収束帯と同じようなはたらきを提供しているように見えます.
しかし,やがて秋がやって来ると,この行き過ぎた北上が,寒さによって,ウスバキトンボを死滅へと追いやるものと思われます.でもここでよく考えてみると,秋になったときには,秋雨前線という収束帯が南日本に生じます.北の方からはこの収束帯に向かって北風が吹きます.シベリア高気圧の勢力が強くなるとともに秋雨前線は南下し,北からの季節風が強く吹くようになります.これに乗れば,ウスバキトンボは南へ帰ることができます.南方定点
Tango での調査でも,9月になると,北西の風に乗って少数のウスバキトンボが現れることが記録されています.ただし寒さに弱いウスバキトンボは,冷涼な北風に乗って,しかも高高度をどれだけ飛べるかということを考えると,この可能性はかなり低くなると思われます.結局,微妙なタイミングで暖かいうちに北風に乗れたごく少数の個体だけが,南への復路移住に成功するのではないかと,私は考えています.
このように,日本にやって来るウスバキトンボは,おそらくほとんどが淘汰される存在になっています.しかし,もし地球温暖化によって,秋の気温低下が遅れたりするようなことになると,この復路移住の成功率が上がってくるかもしれません.ウスバキトンボは,日本においては,北への移住やその出現時期がよく話題になっていますが,南への移動についても,もっと目を向ける必要があるかもしれません.
そういう意味では,八重山諸島のウスバキトンボは,興味ある存在だと思います.石垣島では1月でもウスバキトンボの羽化が見られ,成虫は一年を通じて観察できるので(博物館友の会,1986),気温は問題になりません.八重山諸島は,図11から分かるように,熱帯収束帯が最も北寄りに位置する7月ごろには,その影響下に入っています.そのせいか,石垣島では一年中で6,7月ごろに最も曇りの日が多くなり,70-80%以上の確率で曇ります(Weather Spark,2022.4.25.現在).降水量は7月はやや少ないものの,6-10月にかけて一年で最も多くなります.また卓越する地上の風向は,5-8月は南から南南西で,9月ごろから北北東に変わり,以降それが続きます(以上気象庁,2022.4.25.現在).時期はずれていますが,上記の西インド洋と似たところがあります.
八重山諸島で夏に羽化したウスバキトンボが,その後どの方向に移動していくかといった問題に関しては,いまのところほとんど調査されていないと思われます.今までの議論をもとにすると南への移動が考えられるので,北への移動ばかりが注目されている日本の現状を見たとき,そこには面白い研究テーマが存在するように思います.同じように7月ごろに熱帯収束帯が位置する渤海(中国黄海の奥,図11参照)の研究では,南への移動が示唆されています(
Feng, 2006).
*2.ここでの進化とは小進化のことです.つまり個体群のある個体に生じた突然変異(DNAの塩基配列の変化)が,生殖活動を通じて子孫に伝わりながらやがて集団全体に広がり,ついにはその集団全体がその突然変異を有するような状態になる場合をいいます.したがって,生殖的に隔離された個体群間では,一方に生じた突然変異が他方に伝わらないため,隔離された個体群ごとにわずかに異なる独特のDNAの塩基配列(ハプロタイプ)を持つようになります.
なおこの場合の突然変異(DNAの塩基配列の変化)が表現型の変化を引き起こさない場合(中立突然変異),これが集団に広がるのは,自然選択ではなく,偶然によります(遺伝的浮動).

ウスバキトンボの肛角部は広がっている.ハネビロトンボでも顕著である.
参考文献
Anderson, R. C., 2009. Do dragonflies migrate across the weatern Indian Ocean?. Journal of Tropical Ecology. 25:347-358.
青木典司,1997.標識調査によるベッコウトンボ成虫の動態について.昆虫と自然 32(7):11-18.
朝比奈正二郎・鶴岡保明,1967.南方定点観測船に飛来した昆虫.Kontyu 33(4):353-360.
朝比奈正二郎・鶴岡保明,1968.南方定点観測船に飛来した昆虫 第2報.Kontyu 36(2):190-202.
朝比奈正二郎・鶴岡保明,1969.南方定点観測船に飛来した昆虫類 第3報.Kontyu 37(3):290-304.
朝比奈正二郎・鶴岡保明,1970.南方定点観測船に飛来した昆虫類 第5報.Kontyu 38(4):318-330.
Azuma, A. & T. Watanabe, 1988. Flight performance of a dragonfly. J. exp. Biol., 137:221-252.
Corbet P.S., 1962. A Biology of Odonata. H. F. & G. Witherby ltd., GB.
Corbet P.S., 1999. Dragonflies Behavior and Ecology of Odonata. Cornell Universitiy Press. New York.
Corbet P.S. & Eda S., 1969. Odonata in southern Ontario, Canada in August 1968. Tombo 12:4-11.
DragonflyPix.com., 2022.4.21. http://www.dragonflypix.com/speciespages/pantala_flavescens_sc_cl.html#ShowMap
Dumont H.J. & Hinnekint B.O.N., 1973.Mass migration in dragonflies, especially in Libellula quadrimaculata L.: a review, a new ecological approach and a new hypothesis. Odonatologica 2:1-20.
Feng, Hong-qiang, Kong-ming Wu, Yun-xia Ni, Deng-fa Cheng, and Yu-Yuan Guo, 2006. Nocturnal migration of dragonflies over the Bohai Sea in northern China. Ecological Entomology 31(5).
*Glick P. A., 1939. "The distribution of insects, spiders, and mites in the air." USDA Tech. Bull. 637:1-151. (in Corbet, 1999)
博物館友の会,1986.沖縄のトンボ.70pp. 沖縄県立博物館,那覇.
橋本 碩・朝比奈正二郎,1969.南方定点観測船に飛来した昆虫類 第4報.Kontyu 37(3):305-319.
Hobson, K. A., R. C. Anderson, D. X. Soto & L. I. Wassenaar, 2012. Isotopic evidencetThat dragonflies (Pantala flavescens) migrating through the Maldives come from the Northern Indian Subcontinent. PLoS One 7(12):1-4.
Hobson, K. A., H. Jinguji, Y Ichikawa, J. W. Kusack, and R. C. Anderson, 2021. Long-Distance Migration of the Globe Skimmer Dragonfly to Japan Revealed Using Stable Hydrogen (δ2H) Isotopes. Environmental Entomology 50(1):247-255.
井上 清,1988.淡路島のトンボ相調査会 第3回 秋.Gracile (39):25-32.
*Kallapur V. L., 1985. Some aspects of structual organaization and metabolic adaptations in the flight muscles of the dragonfly Pantala rlavescens. Peoc. 1st Indian Symp. Odonatol., Madurai:53-61. (in Corbet, 1999)
*Kallapur V. L. and George, C. J., 1973. Fatty acid oxidation by flight muscles of the dragonfly, Pantala flavescens. JIP 19:1035-1040. (in Corbet, 1999)
関西トンボ談話会編,1984.近畿のトンボ.大阪.
気象庁ホームページ,2022.4.22.現在.石垣島 平年値(年・月ごとの値).https://www.data.jma.go.jp/obd/stats/etrn/view/nml_sfc_ym.php?prec_no=91&block_no=47918.
李 承模,2001.韓半島産蜻蛉目昆虫誌.正行社,韓国.
May Michael, 1991. Dragonfly flght: Power requirements at high speed and acceleration. J. exp. Biol., 158:325-342.
三時 輝久・平田 真二,1997.ベッコウトンボの移動習性-山口県の生息地におけるマーキング調査の結果より-.昆虫と自然 32(7):27-32.
野平安芸雄,1960.太平洋上でウスバキトンボを発見.Tombo 3(3/4):30-31.
尾園 暁・川島逸郎・二橋 亮,2012.日本のトンボ.文一総合出版.東京.
Rüppell, G., 1989. Kinematic analysis of symmetrical flight manoeuvres of Odonata. J. exp. Biol., 144:13-42.
Papp, R. P., 1974. Recovery of Anax junius from a glacier in the Sierra Nevada (Odonata: Aeschnidae). Pan.-Pacif. Entomol. 50:67.
*Schaefer, G. W., 1976. Radar observations of insect flight. Symp. Roy. Entomol. Soc. Lond. 7:157-197. (in Corbet, 1999)
Snodgrass, R. E., 1935.Principles of insect morphology. Cornel University Press, London.
*Strauss S., 1985. Monarchs: monarchs of flight [interview with D. L. Gibo]. Globe and Mail, Toronto, 18 Nobember, A12. (in Corbet, 1999)
Tillyard, B. J., 1917. The biology of dragonflies (Odonata or Paraneuroptera). Cambridge University Press, London.
津田 滋,2000.世界のトンボ分布目録.自刊.
Ware, J., M. K. Kohli, C. M. Mendoza, D. Troast, H. Jinguji, K. A. Hobson, G. Sahlen, R. C. Anderson & F. Suhling, 2022. Evidence for widespread gene flow and migration in the Globe Skimmer dragonfly Pantala flavescens. International Journal of Odonatology 2: 43-55.
Williams, C. B., 1958. Insect migration. (長澤純夫 訳.1986.昆虫の渡り.築地書館.)
Wojtusiak, J., 1974. A dragonfly migration in the hugh Hindu Kush (Afghanistan), with a note on high altitude records of Aeshna juncea mongolica Bartenev, and Pantala flavescens (Fabricius) (Anisoptera: Aeshnidae, Libellulidae). Odonatologica 3(2): 137-142.
Weather Spark, 2022.4.25.現在.https://ja.weatherspark.com/y/141107/石垣市、日本における年間の平均的な気候#Sections-Clouds.
山口 晴幸,2008.沖縄水史観(X).隆起環礁の島「南大東島」の誕生と日本最大規模を誇るカルスト湖沼群.水利科学 (302):86-100.